






《生物化学 第11章 RNA的生物合成PPT课件下载》是由用户上传到老师板报网,本为文库资料,大小为4.55 MB,总共有93页,格式为ppt。授权方式为VIP用户下载,成为老师板报网VIP用户马上下载此课件。文件完整,下载后可编辑修改。
- 文库资料
- 93页
- 4.55 MB
- VIP模板
- ppt
- 数字产品不支持退货
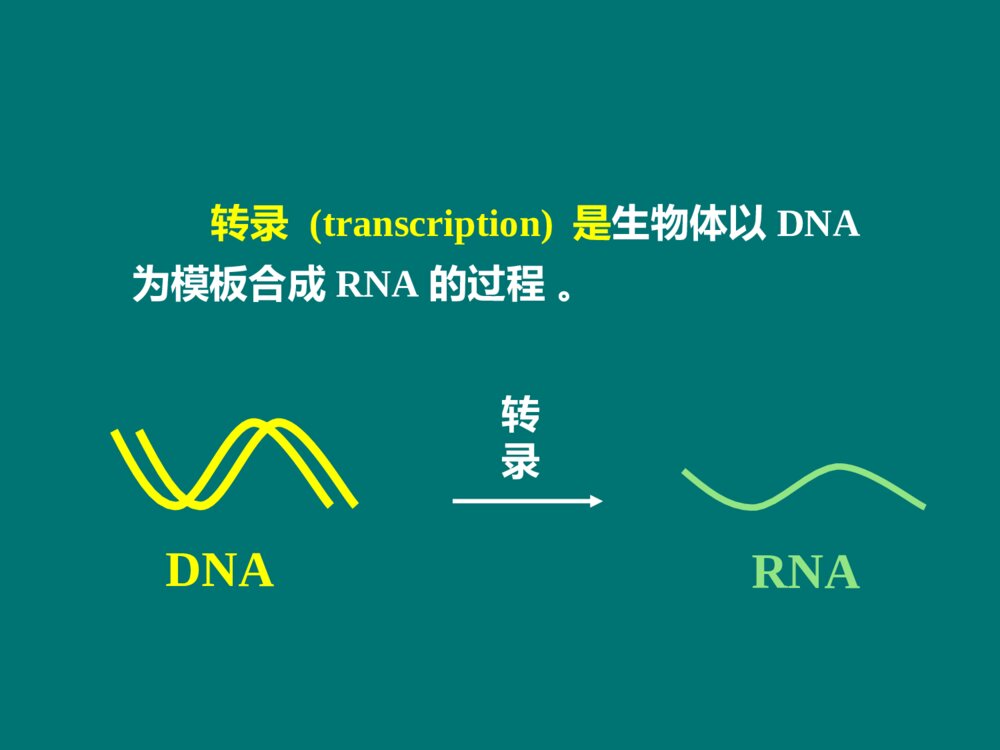
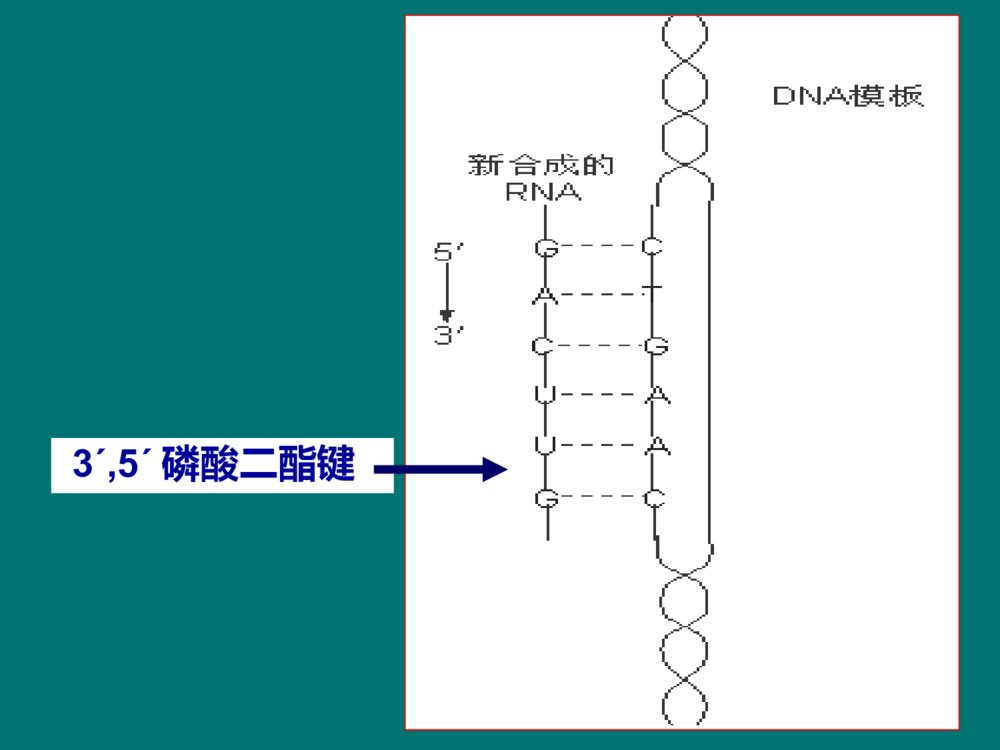
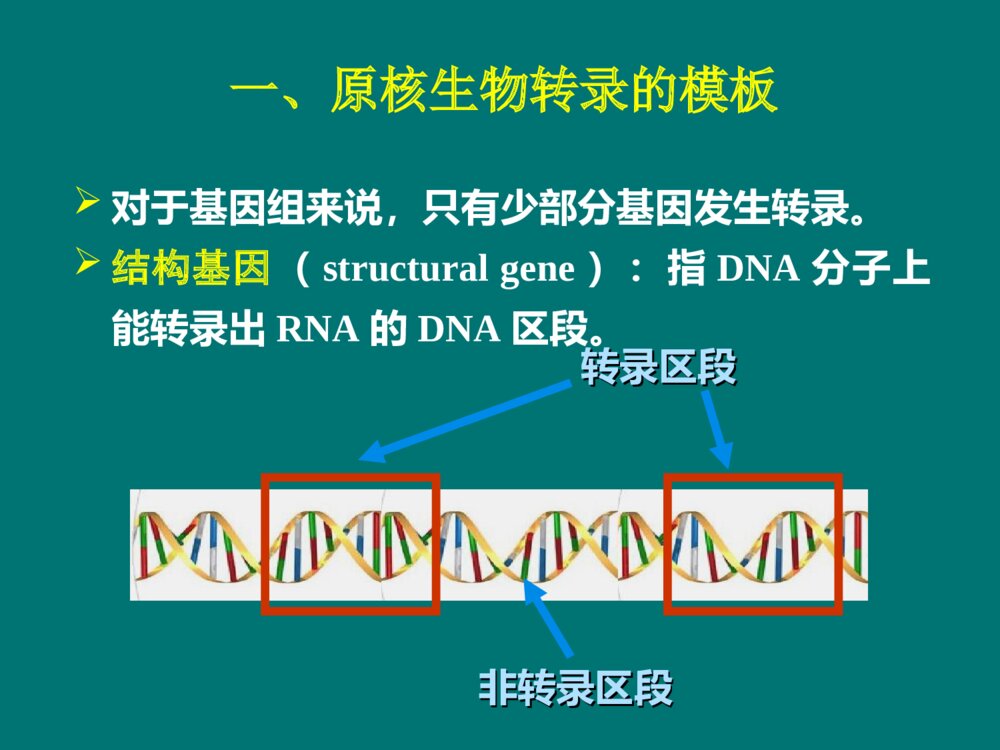
RNABiosynthesis(Transcription)第11章本章内容本章内容原核生物转录的模板和酶(重点)原核生物的转录过程(重点)真核生物的转录过程(一般)真核生物RNA的加工(重点)在生物界,在生物界,RNARNA合成有两种方式:合成有两种方式: 一种是DNA指导的RNA合成,也叫转录,是体内的主要合成方式,也是本章介绍的主要内容。另一种是RNA指导的RNA合成,也叫RNA复制,由RNA依赖的RNA聚合酶催化,常见于病毒,是逆转录病毒以外的RNA病毒在宿主细胞以病毒的单链RNA为模板合成RNA的方式。 概 述转录 (transcription)是生物体以DNA为模板合成RNA的过程 。 转录RNADNA3´,5´磷酸二酯键参与转录的物质参与转录的物质模板:DNA原料:NTP(ATP,UTP,GTP,CTP)酶: 依赖DNA的RNA聚合酶 简称RNA聚合酶或RNA-pol其他蛋白质因子:ρ因子复制与转录的相似点复制与转录的相似点2、都以DNA为模板;1、都是酶促核苷酸聚合反应;3、聚合酶都依赖DNA;4、核苷酸之间都以磷酸二酯键连接;5、合成方向都是5→3;6、都遵从碱基配对规律。复制和转录的区别复制和转录的区别复制转录模板两条链均复制不对称转录原料dNTPNTP酶DNA聚合酶RNA聚合酶引物需要不需要产物两个子代DNA多种RNA配对规律A-T,C-GA-U,T-A,C-G原核生物转录的模板和酶Templates&EnzymesinProkaryoticTranscription第一节一、原核生物转录的模板转录区段转录区段非转录区段非转录区段对于基因组来说,只有少部分基因发生转录。结构基因(structuralgene):指DNA分子上能转录出RNA的DNA区段。5′···GCAGTACATGTC···3′3′···cgtgatgtacag···5′5′···GCAGUACAUGUC···3′N······Ala·Val·His·Val······C编码链(文献所列)模板链mRNA蛋白质转录翻译DNA双链中按碱基配对规律能指引转录生成RNA的一股单链,称为模板链(templatestrand)。相对的另一股单链是编码链(codingstrand)。 5335模板链编码链编码链模板链转录方向转录方向不对称转录(asymmetrictranscription): 转录具有不对称性。3535不对称转录不对称转录有两方面含义:在DNA分子双链上,一股链用作模板指引转录,另一股链不转录;其二是模板链并非总是在同一单链上。5ˊ3ˊ3ˊ5ˊ5ˊ5ˊ5ˊ5ˊ3ˊ3ˊ3ˊ3ˊ二、原核生物RNA聚合酶(一)RNA聚合酶能直接启动RNA链的合成为DNA依赖的RNA聚合酶;聚合反应机制与DNA-pol催化的DNA合成相似;在核苷酸3’-OH上延长RNA链,形成3´,5´磷酸二酯键。(NMP)n+NTP→(NMP)n+1+PPiRNA聚合酶的激活剂:Mg2+和Mn2+5335结构基因调控序列启动子RNA聚合酶不需要引物能直接启动RNA合成。RNA聚合酶和DNA的特殊序列结合后,才能启动RNA合成。 启动子(promoter):是在转录起始上游RNA聚合酶特异性识别和结合的DNA序列。RNA-pol(二)RNA聚合酶由多个亚基组成36512决定哪些基因被转录150618催化功能155613结合DNA模板70263辨认起始点亚基分子量功能ω?功能不详大肠杆菌RNA聚合酶(480kD)组分核心酶( α2ββ′)(coreenzyme)全酶(α2ββ′σ)(holoenzyme)转录起始阶段转录延长阶段 RNA聚合酶的结构示意σ70--辨认典型转录起始点的蛋白质 σ32--辨认热休克蛋白转录起始点的蛋白质 热休克蛋白(Heatshockproteins,Hsp)是细胞在一些应激条件(热休克、葡萄糖饥饿或受到病原菌感染)下有高效表达的一族蛋白。普遍存在于原核和真核细胞中。σ亚基有多种原核生物的RNA聚合酶都受一种抗生素的特异性抑制,即利福平或利福霉素。 利福平或利福霉素是抗结核杆菌的有效药物,机制是专一性地结合RNA聚合酶的 β亚基。原核RNA聚合酶的抑制剂三、三、RNARNA聚合酶与启动子结合启动转录聚合酶与启动子结合启动转录转录是不连续、分区段进行的;每个区段为一个单位。操纵子(operon):是原核转录的功能单位,包括一组结构基因及其上游的调控序列。5335结构基因调控序列启动子启动子是RNA聚合酶结合模板DNA的部位,也是控制转录的关键部位。RNA聚合酶全酶结合到启动子上,其中由σ亚基辨认启动子,其他亚基相互配合。 对启动子的研究,常采用一种巧妙的方法,即RNA聚合酶保护法。5335结构基因调控序列启动子RNA-polσRNA聚合酶保护法40-60bp被RNA聚合酶保护的DNA区段碱基序列分析trpTTGACA•••N17•••TTAACT•••N7•••A•••-35区 -10区tRNAtrpTTTACA•••N16•••TATGAT•••N7•••A•••lacTTTACA•••N17•••TATGTT•••N6•••A•••recATTGATA•••N16•••TATAAT•••N7•••A•••araCTGACG•••N18•••TACTGT•••N6•••A•••最大一致性TTGACATATAAT( Pribnow盒 )+1(转录起始)富含AT开始转录TTGACAAACTGT-35区(Pribnowbox)TATAATATATTA-10区1-30-5010-10-40-205335σ亚基辨认位点(recognitionsite)55RNA聚合酶保护区结构基因33转录起始区RNApol全酶结合部位RNA聚合酶全酶在转录起始区的结合:原核生物的转录过程TheProcessofTranscriptioninProkaryote第二节转录的延长转录的起始转录的终止RNA聚合酶必须准确地结合在转录模板的起始区域。DNA双链解开,使其中的一条链作为转录的模板。一、原核的转录起始转录起始需解决两个问题:E.coliE.coli的转录起始的转录起始2.DNA双链局部(–10区)解开形成开放转录复合体1.σ亚基辨认启动子的识别位点(–35区),RNA聚合酶全酶(2)与启动子结合(–10区),形成闭合转录复合体;3.在RNA聚合酶作用下发生第1个聚合反应,形成转录起始复合物:转录起始过程:4.第1个磷酸二酯键生成后,σ亚基即从转录起始复合物上脱落,核心酶连同四磷酸二核苷酸,继续结合于DNA模板上,酶沿DNA链前移,进入延长阶段;5pppG一直保留。 RNA-pol(2)-DNA-pppGpppGpN-OH3转录起始复合物:5-pppG-OH+NTP5-pppGpN-OH3+ppi目录σ亚基辨认(-35)全酶结合(-10)DNA解链(-10)转录起始复合物形成:2-DNA-pppGpN-OHσ亚基脱落原核转录起始的主要事件:二、原核的转录延长1.亚基脱落,RNA–pol核心酶变构,与模板结合松弛,沿着DNA模板前移;2.在核心酶作用下,NTP不断聚合,RNA链不断延长。(NMP)n+NTP(NMP)n+1+PPi3.延长过程中,酶-DNA-RNA形成转录空泡;转录延长与翻译同时进行。转录空泡(transcriptionbubble):转录延长时,DNA局部双链解开,产物RNA与DNA形成杂化双链,此时酶-DNA-RNA形成的转录复合物被称为转录空泡。G≡C﹥A=T﹥A=U;RNA链5端伸出空泡不断延长5-pppGRNA12bp17bp40bp53DNA原核生物转录过程中的羽毛状现象(电镜下)核糖体RNARNA聚合酶在同一DNA模板上,有多个转录同时在进行;转录尚未完成,翻译已在进行。 这种形状说明:目录核心酶沿模板前移形成转录空泡与翻译同步原核转录延长的主要事件:依赖ρ因子的转录终止非依赖ρ因子的转录终止三、原核生物的转录终止转录终止转录终止指RNA聚合酶在DNA模板上停顿下来不再前进,转录产物RNA链从转录复合物上脱落下来。 依据是否需要蛋白质因子的参与,原核生物转录终止分为:ρ因子是由相同亚基组成的六聚体蛋白质,亚基分子量46kD。ρ因子能结合RNA,又以对polyC的结合力最强。ρ因子还有ATP酶活性和解螺旋酶的活性。(一)依赖ρ因子的转录终止ρ因子的特点:ρ因子的作用原理:ρ因子与RNA转录产物(3’端富含C)结合后,ρ因子和RNA聚合酶构象变化,使RNA聚合酶停顿,解螺旋酶的活性使DNA/RNA杂化双链拆离,利于产物从转录复合物中释放 。(二)非依赖ρ因子的转录终止DNA模板上靠近转录终止处,有些特殊的碱基序列,转录出RNA后,RNA产物形成特殊的结构来终止转录。近终止区的转录产物形成茎环或发夹结构是非依赖ρ因子终止的普遍现象。 茎环结构使转录终止的机理:茎环结构使RNA聚合酶变构,转录停顿;茎环结构使转录复合物趋于解离。杂化双链不稳定(rU/dA最不稳定);寡聚U促使RNA脱落。5´pppG5335RNA-pol目录依赖ρ因子的转录终止:ρ因子结合RNA,杂化双链解开,RNA释放。不依赖ρ因子的转录终止:RNA形成茎环结构,杂化双链不稳定,RNA释放。原核转录终止的主要事件:小结1、概念:不对称转录、模板链、编码链、启动子、因子、转录空泡2、原核RNA聚合酶的亚基组成及功能3、原核生物转录的过程:起始、延长、终止思考题1、试比较复制与转录的异同点。2、简述原核生物的转录过程。真核生物的转录过程TheProcessofTranscriptioninEukaryote第三节真核生物的转录过程比原核复杂。RNA聚合酶种类不同,与模板结合的特性也不同;转录起始过程有较大区别,转录延长和终止也不相同。 一、真核生物有三种DNA依赖性RNA聚合酶真核生物RNA聚合酶的种类:RNA聚合酶Ⅰ(RNAPolⅠ)RNA聚合酶Ⅱ(RNAPolⅡ)RNA聚合酶Ⅲ(RNAPolⅢ)真核生物的RNA聚合酶种类ⅠⅡⅢ位置核仁核质核质转录产物45s-rRNAhnRNAsnRNA5s-rRNAtRNAsnRNA对α鹅膏蕈碱的反应耐受极敏感中度敏感45s-rRNA加工成28s、5.8s、18srRNA真核还有线粒体、叶绿体RNA聚合酶 真核生物RNA聚合酶的结构:大亚基:2个(与原核的βˊ、β相似)小亚基:十几个RNA聚合酶Ⅱ有12个亚基。最大亚基RBP1羧基末端有Tyr-Ser-Pro-Thr-Ser-Pro-Ser重复序列,称为羧基末端结构域(CTD)。 所有RNA聚合酶Ⅱ都有CTD,只是重复程度不同(哺乳动物52次,酵母菌26-29次,果蝇44次);去磷酸化CTD参与转录起始,磷酸化CTD参与延长。RNA聚合酶Ⅰ、Ⅲ没有CTD。二、真核转录起始需要启动子、二、真核转录起始需要启动子、RNARNA聚合酶和转录因子的参与聚合酶和转录因子的参与(一)转录起始前的上游区段具有启动子核心序列不同基因转录起始点上游存在有各自的调控序列,即顺式作用元件(cis-actingelement)。顺式作用元件是指影响自身基因表达活性的DNA序列,包括启动子、启动子上游元件、增强子、沉默子等。启动子的核心序列: 起始点上游-30处多数有共同的TATA序列,称为Hogness盒或TATA盒(TATAbox)。 位于转录起始点附近的起始子(intiator,Inr),富含嘧啶序列;也是启动子的保守序列 。 3’5’上游元件TATA盒Inr起始子YYANYYTA-30+1TATAAA启动子上游元件:位于TATA盒上游,多在约-40~-100nt的位置,比较常见的是GC盒和CAAT盒。增强子:是能够结合特异基因调节蛋白, 促进邻近或远隔特定基因表达的DNA序列,以上游为主。(二)转录因子转录因子(transcriptionfactors,TF)是一类能与特异性DNA序列结合并调节基因转录的蛋白质。基本转录因子:RNA聚合酶启动基因转录时所必需的一类蛋白质,协助RNA聚合酶与启动子结合并起始转录。RNApolⅠ、Ⅱ、Ⅲ的TF,称为TFⅠ、TFⅡ、TFⅢ;各有亚类,在进化中高度保守。参与RNA-polⅡ转录的TFⅡ*TBP:TATA结合蛋白 ;**TAF:TBP辅助因子,人类至少有12种TAF。不同TAF与TBP结合可结合不同的启动子。蛋白激酶活性,使CTD磷酸化TFⅡHATPase57()34()TFⅡE解螺旋酶30,74TFⅡF促进RNA-polⅡ结合及作为其他因子结合的桥梁33TFⅡB稳定TFⅡD-DNA复合物12,19,35TFⅡA辅助TBP-DNA结合TAF**结合TATA盒TBP*38TFⅡD功能亚基组成,分子量(kD)转录因子蛋白激酶活性,使CTD磷酸化TFⅡHATPase57()34()TFⅡE解螺旋酶30,74TFⅡF促进RNA-polⅡ结合及作为其他因子结合的桥梁33TFⅡB稳定TFⅡD-DNA复合物12,19,35TFⅡA辅助TBP-DNA结合TAF**结合TATA盒TBP*38TFⅡD功能亚基组成,分子量(kD)转录因子;解螺旋酶ⅡⅡ型基因中的四类转录因子型基因中的四类转录因子转录因子 具体组分 结合序列 功能 基本转录因子 TBP,TFA,ⅡB,E,F和HTBP结合TATA盒 转录起始定位; 转录起始和延长 辅激活因子 TAFs和中介子 在可诱导因子和上游因子与基本转录因子、RNA聚合酶结合中起联结和中介作用 上游因子 SP1、ATF、CTF等 启动子上游元件 协助基本转录因子,提高转录效率和专一性 可诱导因子 如MyoD、HIF-1等 增强子等远隔序列 时间和空间(组织)特异性地调控转录 RNA聚合酶II与启动子的结合、启动转录需要多种蛋白质因子的协同作用。通常包括:可诱导因子或上游因子与增强子或启动子上游元件的结合;转录因子在启动子处的组装;辅激活因子和/或中介子在转录因子/RNA聚合酶II复合物与可诱导因子、上游因子之间的辅助和中介作用。 因子和因子之间互相辨认、结合,以准确地控制基因是否转录、何时转录。(三)转录起始前复合物真核生物RNA-pol不与DNA分子直接结合,而需依靠众多的转录因子,形成转录起始前复合物(pre-initiationcomplex,PIC): PIC是一个闭合复合体:DNA(TATA)-TBP-A-B-F-E-H-RNA-polⅡPIC的形成TFDⅡ的TBP结合TATA盒;TFBⅡ与TBP、DNA结合(TFAⅡ配合);TFFⅡ与RNApol结合两个复合体结合,TFFⅡ与TFBⅡ相互作用,协助RNApol结合启动子TFEⅡ和H加入,形成闭合复合体,PIC形成开放复合体形成,转录起始当合成60-70nt时,TFEⅡ和H释放,转录进入延长期。TFHⅡ(解旋酶)解开DNA双链,启动转录;使RNApol的CTD磷酸化(激酶活性)。(四)少数几个转录因子的搭配启动特定基因(四)少数几个转录因子的搭配启动特定基因的转录的转录为了保证转录的准确性,不同基因需不同转录因子。发现的TF不断增加。拼板理论:转录因子之间有不同的搭配 少数几个反式作用因子之间互相作用,再与基本转录因子、RNA聚合酶搭配而有针对性地结合、转录相应的基因。估算300多个转录因子即能满足需要。真核生物转录起始的特点真核生物转录起始的特点转录起始上游区段的调控序列更加复杂:包括启动子、启动子上游序列、增强子等RNA聚合酶和模板的结合更加复杂:不直接结合模板,需要众多转录因子的参与。三、真核生物转录延长过程中没有转录与翻译同步的现象真核生物转录延长过程与原核生物大致相似,但因有核膜相隔,没有转录与翻译同步的现象。 RNA-pol前移过程中处处都遇上核小体。转录延长过程中可以观察到核小体移位和解聚现象。 RNA-PolRNA-PolRNA-Pol核小体转录延长中的核小体移位转录方向四、真核生物的转录终止和加尾修饰同时进行真核生物mRNA有聚腺苷酸(polyA)尾巴结构,是转录后才加进去的。转录不是在polyA的位置上终止,而是超出数百个乃至上千个核苷酸后才停顿。已发现,在读码框架的下游,常有一组共同序列AATAAA,再下游还有相当多的GT序列。这些序列称为转录终止的修饰点。5’------AAUAAA-5’------AAUAAA—Poly(A)RNA核酸酶-GUGUGUGAATAAAGTGTGTG转录终止的修饰点5’5’3’3’RNA-pol真核生物的转录终止AAAAA…AAAAA真核生物RNA的加工Post-transcriptionalModificationofEukaryoticRNA第四节真核生物转录生成的RNA分子是初级RNA转录物,几乎所有的初级RNA转录物都要经过加工,才能成为具有功能的成熟的RNA。加工主要在细胞核中进行。一、真核生物mRNA的加工包括首、尾修饰和剪接(一)前体mRNA在5’-末端加入“帽”结构前体mRNA也称初级mRNA转录产物,或不均一核RNA(hnRNA)。大多数真核mRNA的5’-末端有帽子结构。11、帽子结构:、帽子结构:7-7-甲基鸟苷三磷酸(甲基鸟苷三磷酸(m7Gppp-m7Gppp-))5pppGp…5GpppGp…pppGppi加帽酶5m7GpppGp…甲基转移酶SAM2、帽子结构的生成过程:5ppGp…加帽酶Pi55-三磷酸结构目录HH帽子0型:m7GpppNp帽子1型:m7GpppNmpNp帽子2型:m7GpppNmpNmpNp33、帽子结构的意义:、帽子结构的意义:可以保护mRNA免遭核酸酶的攻击;与帽结合蛋白质复合体结合,参与mRNA和核糖体的结合,启动蛋白质的生物合成。 真核生物成熟mRNA(除组蛋白), 3’端都有polyA结构,约为80至250个核苷酸之间。多聚腺苷酸尾的生成是多步骤过程,与转录终止同时进行。3’多聚腺苷酸化的过程如下:(二)前体mRNA在3’端特异位点断裂并加上多聚腺苷酸尾AAUAAAG/U5’3’PolyA信号PolyA位点mRNA断裂与聚腺苷酸化特异性因子(CPSF)G/U5’3’CPSF断裂因子CFI,CFII,断裂激动因子CStFCPSFCStFCFICFII多聚腺苷酸聚合酶(PAP)CPSFCStFCFICFIIPAPATPPPiCFIICFICStF多聚腺苷酸结合蛋白Ⅱ(PABⅡ)CPSFAAAAAAAAAAOHPAPATPPPiPABⅡ快速多腺苷酸化,在足够长时终止CPSFAAAAAAAAAAAAAAAAAAAAHOAAAAAAAAAAPAPPAP慢速多腺苷酸化CPSFAAAAAAAAAAOH形成CPSF-信号复合体CFI,CFII,CStF结合CPSF-信号复合体G/UP3’端polyA的意义polyA维持mRNA作为翻译模板的活性,增加mRNA本身稳定性。序列分析证明mRNA来源于hnRNA,只是去除了大部分中间片断。杂交实验证明hnRNA与DNA模板链完全互补;mRNA与DNA杂交,出现配对双链区和鼓泡的单链区1.hnRNA和断裂基因(splitegene)(三)前体(三)前体mRNAmRNA的剪接的剪接DNAmRNA真核生物结构基因,由若干个编码区和非编码区互相间隔排列,去除非编码区再连接后,可翻译出由连续氨基酸组成的完整蛋白质,这些基因称为断裂基因。 断裂基因(splitegene)CABD编码区 A、B、C、D非编码区2.外显子(exon)和内含子(intron)外显子:在断裂基因及其初级转录产物上出现,并表达为成熟RNA的核酸序列。内含子:隔断基因的线性表达而在剪接过程中被除去的核酸序列。 少数真核基因无内含子,如组蛋白基因、低等真核生物(酿酒酵母)。广义的内含子包括翻译后删除内含子(胰岛素原的C肽)。 最庞大的抗肌萎缩蛋白基因(106bp)分别有50多个外显子和内含子;内含子的功能?鸡卵清蛋白基因hnRNA首、尾修饰hnRNA剪接成熟的mRNA鸡卵清蛋白基因及其转录、转录后修饰套索RNA外显子内含子鸡卵清蛋白成熟mRNA与DNA杂交电镜图DNA(7.7kb)mRNA(1.2kb)3.mRNA的剪接--指去除hnRNA中的内含子,将外显子连接为成熟的mRNA,称为剪接。剪接接口:5’GU……AG3’剪接的场所--剪接体(超大分子复合体) 剪接体:由多种小分子核糖核蛋白(snRNP)组成snRNP:是snRNA-蛋白质复合体,其RNA和蛋白质都高度保守。snRNA有5种:U1、U2、U4、U5、U6,长度100-300nt,富含尿嘧啶。mRNA的剪接机制E1E2①②③UACUACA-AGUGU4U5U6E1E2U1U2UACUACA-AGUGU6E1E2U1、U4、U55′GU….AG3′为剪接接口U2U2、U6形成催化中心U1、U2与内含子两端配对; U4、U5、 U6加入;内含子弯曲成套索,E1、E2靠近pG-OH(ppG-OH,pppG-OH)G-OHApGpGpU第一次转酯反应第二次转酯反应GpUApG外显子1内含子外显子2A-OHGpGpGpU•剪接过程的二次转酯反应(twicetransesterification)剪切--剪去内含子后不连接(位于末端)剪接--剪去内含子后把外显子连接起来 经过选择性剪接可以产生不同的mRNA4.前体mRNA经剪接和剪切两种模式可加工成不同的mRNA•RNA编辑作用说明,基因的编码序列经过转录后加工,是可有多用途分化的,因此也称为分化加工(differentialRNAprocessing)。(四)mRNA编辑(mRNAediting)人类apoB基因mRNA(14500个核苷酸)肝脏(2153Gln,CAA)apoB100(分子量为500000)肠道细胞(UAA)apoB48(分子量为240000)C→U--在mRNA中插入、缺失或替换核苷酸而改变DNA模板来源的遗传信息 。二、前体rRNA的转录后加工转录45S-rRNA剪接18S-rRNA5.8S和28S-rRNArDNA内含子内含子28S5.8S18SRNApolⅠ三、前体tRNA的转录后加工tRNA前体RNApolⅢTGGCNNAGTGCGGTTCGANNCCDNARNAaseP、RNAaseD、内切酶5’端核苷酸序列切除;3’端UU切除;切除内含子。核苷酸转移酶ATPADP3’加上CCA碱基修饰(2)还原反应 如:UDHU(3)核苷内的转位反应 如:Uψ(4)脱氨反应 如:AI 如:AAm(1)甲基化(1)(1)(3)(2)(4)四、RNA催化一些内含子的自剪接1982年美国科学家T.Cech发现四膜虫rRNA前体能准确地自我剪接去除内含子。后来在其他rRNA、tRNA、mRNA前体也发现了此类自我剪接的内含子。即RNA有催化作用,称为核酶。自剪接--RNA催化自身内含子剪接的反应组Ⅰ内含子:以鸟嘌呤核苷酸为辅因子,是游离的。组Ⅱ内含子:类似mRNA的剪接,但不需剪接体。小结1、真核RNA聚合酶的种类、特点2、真核生物基因转录的特点3、真核3种RNA加工的主要特点4、概念:帽子结构、断裂基因、外显子、内含子思考题试比较原核与真核生物转录的区别。真核生物转录的特点真核生物转录的特点复杂酶不同;与模板结合的特性不同;TATA盒、起始子;需要转录因子细胞核进行;没有转录与翻译同步的现象有核小体移位现象终止与加尾同时进行mRNA为单顺反子;产物加工修饰复杂:剪接内含子等
 九年级化学下册·初三《《化学肥料》(第11.2课时)PPT课件
九年级化学下册·初三《《化学肥料》(第11.2课时)PPT课件 电镜细胞化学技术PPT课件下载(共56页)
电镜细胞化学技术PPT课件下载(共56页) 九年级化学《第八单元 第四节 常用的盐 课题 盐的化学通性》PPT课件下载
九年级化学《第八单元 第四节 常用的盐 课题 盐的化学通性》PPT课件下载 高中化学干燥剂小结PPT课件下载
高中化学干燥剂小结PPT课件下载 高中化学《第一节 有机化合物的分类》PPT课件下载
高中化学《第一节 有机化合物的分类》PPT课件下载 粤教版化学九年级下册6.4《珍惜和保护金属资源》PPT课件下载
粤教版化学九年级下册6.4《珍惜和保护金属资源》PPT课件下载